Review
| Rev Diabet Stud,
2016,
13(2-3):113-131 |
DOI 10.1900/RDS.2016.13.113 |
The Physiological Effects of Dandelion (Taraxacum Officinale) in Type 2 Diabetes
Fonyuy E. Wirngo, Max N. Lambert, Per B. Jeppesen
Department of Endocrinology and Internal Medicine, Aarhus University Hospital, Tage-Hansens Gade 2, DK-8000 C, Denmark
Address correspondence to: Per B. Jeppesen, e-mail: per.bendix.jeppesen@clin.au.dk
Manuscript submitted October 6, 2015; resubmitted November 6, 2015; accepted December 14, 2015.
Keywords: type 2 diabetes, dandelion, chlorogenic acid, chicory acid, taraxasterol, sesquiterpene
Abstract
The tremendous rise in the economic burden of type 2 diabetes (T2D) has prompted a search for alternative and less expensive medicines. Dandelion offers a compelling profile of bioactive components with potential anti-diabetic properties. The Taraxacum genus from the Asteraceae family is found in the temperate zone of the Northern hemisphere. It is available in several areas around the world. In many countries, it is used as food and in some countries as therapeutics for the control and treatment of T2D. The anti-diabetic properties of dandelion are attributed to bioactive chemical components; these include chicoric acid, taraxasterol (TS), chlorogenic acid, and sesquiterpene lactones. Studies have outlined the useful pharmacological profile of dandelion for the treatment of an array of diseases, although little attention has been paid to the effects of its bioactive components on T2D to date. This review recapitulates previous work on dandelion and its potential for the treatment and prevention of T2D, highlighting its anti-diabetic properties, the structures of its chemical components, and their potential mechanisms of action in T2D. Although initial research appears promising, data on the cellular impact of dandelion are limited, necessitating further work on clonal β-cell lines (INS-1E), α-cell lines, and human skeletal cell lines for better identification of the active components that could be of use in the control and treatment of T2D. In fact, extensive in-vitro, in-vivo, and clinical research is required to investigate further the pharmacological, physiological, and biochemical mechanisms underlying the effects of dandelion-derived compounds on T2D.
Abbreviations: ADP - adenosine diphosphate; AFLD - alcoholic fatty liver disease; AMPK - adenosine monophosphate-activated protein kinase; ATP - adenosine triphosphate; cAMP - cyclic adenosine monophosphate; CGA - chlorogenic acid; CoA - coenzyme A; CRA - chicory acid; DAG - diacylglycerol; DBD - DNA-binding domain; DNA - deoxyribonucleic acid; DPPH - 2,2-diphenyl-1-picrylhydrazyl; Dw - dry weight; FOS - fructose oligosaccharide; G6P - glucose-6-phosphate; GDP - guanosine 5'-diphosphate; GLP-1 - glucagon-like peptide 1; GLUT2 - glucose transporter 2; GLUT4 - muscle glucose transporter protein 4; GPCR - G protein-coupled receptor; GTP - guanosine triphosphate; HNB - 2-hydroxy-5-nitrobenzenaledehyde; HPLC - high-pressure liquid chromatography; IC50 - half maximal inhibitory concentration; IDF - International Diabetes Federation; IDX-1 - islet duodenum homeobox 1; IL-1α - interleukin 1 alpha; INS-1E - rat insulinoma clonal beta-cell line; IR - insulin receptor; IRS-1 - insulin receptor substrate 1; Km - Michaelis constant; IP3 - inositol triphosphate; IRS-1 - insulin receptor substrate 1; LBD - ligand-binding domain; LC-DAD - liquid chromatography with (photo) diode array detection; LPS - lipopolysaccharide; MAPK - mitogen-activated protein kinase; NADH - nicotinamide adenine dinucleotide; NAFLD - non-alcoholic fatty liver disease; NF-κb - nuclear factor kappa B; NO - nitric oxide; PI3K - phosphatidylinositol 3 kinase; PKA - protein kinase A; PKC - protein kinase C; PPAR-γ - peroxisome proliferator-activated receptor gamma; ROS - reactive oxygen species; RxR - retinoid X receptor; SEL - sesquiterpene lactones; SUR1 - sulphonylurea receptor 1; T2D - type 2 diabetes; TAG - triacylglycerol; TNF-α - tumor necrosis factor; TO - Taraxacum officinale; TS - taraxasterol; UPLC-MS/MS - ultra-performance liquid chromatography - tandem mass spectrometry; UV/VIS - ultraviolet visible; WHO - World Health Organization
1. Introduction
Societies in both developed and developing countries are engulfed by the metabolic disorder of type 2 diabetes (T2D). The world is facing a huge clinical and economic burden due to the enormous increase in diabetes incidence. It is estimated that approximately 382 million people in the world have T2D today, and by 2035, this number is expected to rise by more than 200 million if preventive measures are not established [1]. A WHO survey indicated that 70-80% of the world’s population is relying on non-conventional medicines, primarily because of a lack of availability of and economic barriers to conventional medicine. In the past, plant-derived therapeutics have been widely disregarded as a possible cost-effective means to treat diabetes; hence evidence-based documentation of efficacy is commonly unavailable. In spite of this deficit, it is well known that plant-derived therapeutics provide promising sources of alternative treatment measures, which can even lead to improved efficacy and reduced side effects in comparison to existing conventional medicines [2]. Therefore, there has been increasing interest in food, nutraceuticals, and medicinal products from plants and other natural sources that retain beneficial health properties in developed countries [3].
According to statistics from the International Diabetes Federation (IDF), 80% of people with T2D live in countries characterized by low and middle income. Even more alarmingly, it is estimated that 175 million people with diabetes still go undiagnosed [4]. In poorer regions, treatment of diabetes is very expensive, which makes medical treatment unattainable, resulting in poor healthcare and the use of alternative medicine [5]. Traditional medicine involving the use of bioactive plants has demonstrated potential to alleviate diabetic symptoms, enable recovery, and improve health [6]. Diabetes treatment has been attempted with different plants and poly-herbal formulations, with anti-diabetic activities originating from their bioactive components [7]. About 80% of people worldwide use traditional medicine, while approximately 75% of modern pharmaceuticals are derived from plants [8]. Medicinal plants include a wide variety of anti-diabetic components; frequently their discovery arises from ethnomedical knowledge [9, 10].
The metabolic syndrome, characterized by obesity, hypertension, cardiovascular abnormalities, coronary artery disease, and dyslipidemias, is a core feature of T2D. This non-communicable disease is a metabolic disorder that involves alterations in carbohydrate, lipid, and protein metabolism, as well as pancreas function [7, 11]. T2D is a chronic multifactorial disease, resulting from defects in insulin and glucagon secretion and action, which may cause a progressive increase in plasma glucose levels and a disruption of biological mechanisms in liver, endocrine pancreas, skeletal muscle, adipose tissue, central nervous system, and gut, causing the dysregulation of glucose homeostasis, which plays a key role in the development of T2D [12]. T2D is a common endocrine disorder leading to increased water and food consumption, lipid formation, hyperglycemia, and elevated insulin production, which reinforces existing insulin resistance and contributes to pancreatic failure [13-15]. Insensitivity to insulin leads to dysregulation of muscles, fat, and liver cells due to inadequate transportation of glucose and abnormal storage of lipids [16, 17]. Eventually, chronic diabetes can cause blindness and renal failure, and is a major risk factor for cardiovascular diseases and stroke. In severe cases, it may result in lower limb amputations [13].
The aim of this review is to evaluate the properties of a promising herbal candidate, dandelion, and to explore its diverse biological activities relevant to T2D, with a particular focus on the most current literature regarding the effects of its bioactive components on insulin function and glucose homeostasis.
2. Origin and botany of dandelion
Dandelion is a plant of the genus Taraxacum and a member of the Asteraceae family. It has been used as a medicinal herb for a long time. Figure 1 shows an image of the dandelion plant including its various components that may be used for formulations [18]. Dandelion is produced for medicinal purposes and food, either grown from wild sources or cultivated. It is predominantly cultivated and produced in Bulgaria, Romania, Hungary, and Poland [19]. Dandelion occurs in the tropics, in cool highlands (1,200-1,500 m of altitude) and in warm sub-temperate and temperate zones around the northern hemisphere. It is able to tolerate drought and frost [20, 21].
 |
 |
Figure 1. Image of dandelion and some components present at the level of flowers, stems, and leaves. |
|
Taraxacum is taxonomically complex. In subarctic and Northern temperate regions, there are around 2800 known species [22]. The species Taraxacum officinale WEBER WIGG originates from Europe, and is used for medicinal purposes, while Taraxacum platycarpum is used as a Chinese traditional medicine to treat T2D and hepatic diseases [23]. As a perennial weed, dandelion produces a stout taproot with an average length of 15-30 cm [24]. Even if dandelion is cut below soil surface, the remaining roots are able to generate new plants. On average, each plant develops 5 to 10 flowers containing brown, conical fruits with a hairy pappus, which allows seeds to be distributed by wind. Young dandelion plants are cultivated under glass, and bedded out into manure soil. They are often sown between April and June in northern Europe [25].
In Russia, India, and China, dandelion has been used in ethnopharmacology as a traditional folk medicine because of its hepatic and hyperglycemic effects [8]. It is often consumed as a food (salads) as it is a rich source of micronutrients such as minerals and vitamins [26]. Dandelion has numerous therapeutic benefits, including treatment of T2D, blisters, spleen, and liver complaints [27], and is used as a popular traditional medicine in Turkey and Mexico for the control of T2D [28].
3. Nutritional, chemical, and biological properties of dandelion
Tandem liquid chromatography and mass spectrometry analysis of dandelion have revealed rich sources of β-carotene, which protects cells from oxidation and cellular damage [29]. Recently, biochemical analysis identified chicoric acid (CRA) as the most abundant component of dandelion, with a polyphenolic amount of 34.08 ± 1.65 g/kg in Taraxacum officinale leaves and stems [30-32]. The concentration of polyphenols is higher in flowers and leaves than in stems [33]. There is also considerable seasonal variation; methylsterols are highly present in the winter, while sitosterol and cycloartenol esters are more prevalent during periods of extensive sun exposure [34].
The roots of dandelion contain carbohydrates (e.g. inulin), carotenoids (e.g. lutein), fatty acids (e.g. myristic acid), minerals, sugars (e.g. glucose, fructose, and sucrose), choline vitamins, mucilage, and pectin. Up to 45% of the roots consist of inulin, a complex carbohydrate (fructo-oligosaccharides) with many beneficial effects such as the elimination of pathogens in the gastrointestinal tract, and repression of obesity, cancer, and osteoporosis [35]. Besides its use as a coffee substitute and flavor enhancer in drinks, the leaf extracts are known to be effective against obesity and cardiovascular disease [36].
Dandelion includes potential bioactive components such as sesquiterpene lactones, taraxasterol (TS), taraxerol, chlorogenic acid (CGA), and CRA. These components are non-toxic and can be exploited for their potentially anti-inflammatory, anti-oxidative, anti-rheumatic, and chloretic properties [37]. See Table 1 for an overview of names and structures of dandelion components with antidiabetic actions. To date, there is little research investigating the content of the interesting dandelion components, in particular TS and taraxerol. However, dandelion has been shown to have a high content of these components compared to other plants; it contains a broad range of interesting bioactive components relevant to T2D that are likely to exert synergistic actions [38].
Table
1.
Names and structures of dandelion components with antidiabetic actions (Chembiodraw ultra 14.0) |
|
 |
|
3.1 Sesquiterpene lactones
Sesquiterpene lactones (SEL) are common in plants of the Asteraceae family. The biological profile includes anti-bacterial, fungicidal, growth-regulating, anti-mutagenic, anti-feedant, and repellent properties [39]. Sesquiterpenes are colorless lipophilic components; their biosynthesis is from isoprene units, which occurs through farnesyl pyrophosphate found in endoplasmic recticulum [40]. Sesquiterpenes are generally extracted from the root, and consist of various components, including (Table 1):
- Eudesmanolides tetrahydoridentin B
- Taraxacolide-0-β-glucopyranoside
- 13-dihydrolactucin
- Ixerin D
- Germacranolide acid
- Taraxinic acid
- β-glucopyranoside
- Ainslioside
- 13-dihydrotaraxinic-acid β-glucopyranoside [41]
The bitterness resulting from sesquiterpene lactones is due to components such as taraxacolide, dihydro-lactucin, ixerin D, taraxinic acids, phenyl propanoids, and ainslioside, which have anti-inflammatory properties [42]. These components are often present as glycosides. The bitter taste receptor (TAS2R16) on the cells of the tongue has a protective property to prevent the ingestion of toxic substances, eliciting an aversive sensory response [43]. However, there are flaws to this evolutionary system as a large proportion of substances beneficial to health taste bitter. The health-promoting benefits of dandelion can be attributed to the presence of these bitter substances and of phenolic components, which possess anti-oxidative and anti-inflammatory activities [44]. The anti-inflammatory properties are mediated chemically by the oxygen-containing ring structure featuring a carbonyl function (α-methyl-γ-lactone). This structure reacts with nucleophiles like cysteine sulfhydryl groups [45]. Thiol groups such as cysteine protein residues are known to be the primary target group of sesquiterpene lactones [46]. Sesquiterpene lactones do not exert direct anti-oxidant activity, which is attributable to their structure, but could show greater efficacy as a direct anti-oxidant due to the presence of allylic alcohol [47, 48].
3.2 Taraxasterol
Taraxasterol (TS) is a pentacyclic-triterpene, which is highly present in dandelion roots throughout the year [49]. It is common in esculent plants such as legumes, cereals, nuts, and seeds as well as plant oils [50]. A study using high-pressure liquid chromatography (HPLC) ultraviolet-visible (UV/VIS) detection found high quantities of TS (2.96 µg/ml) and taraxerol (1.69 µg/ml) in natural root extract of Taraxacum Officinale weber; in callus culture, the root extract included 3.0 µg/ml TS and 1.75 µg/ ml taraxerol [38]. It is also extracted from the flowers of Carthamus tinctorius, Chrysanthemum morifolium, and Helianthus annuus. According to Zhang et al. (2012), TS has anti-inflammatory properties, and is considered a therapeutic agent for the treatment of inflammatory diseases. It is known as a monohydroxy triterpene, which is found in burdock, arnica, and chicory, and which displays several biological activities [51]. TS has inhibitory effects on inflammatory ear edema in mice, and it may retain chemo-preventive properties [52]. It is a naturally occurring sterol derived from hydroxylated polycyclic isopentenoid that has the structure of 1,2-cyclopentanophenanthrene. Apart from their importance in agricultural products for the food industry, sterols have a wide range of diverse biological activities, representing an economic value for the pharmaceutical industry [53] TS has shown anti-microbial activity against Staphylococcus Aureus; its presence in cancer cells may counteract the development of tumor at various stages [54].
3.3 Chlorogenic and chicoric acid
CGA is an ester of caffeic and quinic acid and the richest phenolic component of dandelion. It is found in all parts of the plant (roots, flowers, leaves, and stems). A study using ultra-performance liquid chromatography - tandem mass spectrometry (UPLC-MS/MS) showed that the synthetic commercial standard Trolox® is a less effective anti-oxidant than natural CGA. An examination of the phenolic content of a fractionated ethyl acetate extract from dandelion roots showed that CGA was the most common component (31.06 mg/g). Other studies using liquid chromatography with (photo) diode array detection (LC-DAD) indicated that the concentration in leaf extracts amounts to 0.85 mg/g, with 0.22% dry weight (Dw); HPLC-UV analysis showed concentrations to be 8 mg/g in leaf extracts. LC-DAD analysis indicated CRA at 0.09-0.51% Dw in roots and 0.77% Dw in the leaves [55-58]. According to Owen et al. (2014), CGA is a potent anti-oxidant because of its high phenolic content, with an ethyl acetate chromatographic fraction of 2,2-diphenyl-1-picrylhydrazyl (DPPH) IC50 (0.033 ± mg/ml) [55].
CGA consists of major phenolic components which generate the natural anti-oxidant property, possibly by binding to enzymes or multi-subunits of proteins, thereby altering their biological activities [59]. Thus, CGA may be used as an anti-diabetic, anti-oxidant, and anti-carcinogenic agent, which is based on its 5-0-caffeoylquinic acid structure and its scavenger activity for reactive oxygen and nitrogen species [60].
CRA is the most abundant component found in the roots, leaves, and stems of dandelion. This was revealed by the analysis of thirty phenolic components of dandelion juice, using liquid chromatography, mass spectrometry, and HPLC [30]. 2,3-dicaffeoyltartaric acid is a phenolic component from Chichorium intybus, which is known to be a strong anti-diabetic agent [61]. CGA and CRA originate from plants of the same family (Chichorium intybus), and have been used as raw materials for the generation of coffee substitutes [62]. In the presence of inulin, CRA improves lipid metabolism. In an in-vivo study, rats fed with chicory extract and those fed with an inulin-enriched diet showed a significant increase in fecal lipid, cholesterol, and bile acid excretion, implying that chicory regulates lipid metabolism [63].
3.4 Other micronutrients found in dandelion
Dandelion is also rich in vitamins (A, C, D, E, and B), inositol, lecithin, and minerals such as iron, magnesium, sodium, calcium, silicon, copper, phosphorus, zinc, and manganese [64]. The flow of some of these ions, e.g. calcium ions in beta-cells, may help to stimulate insulin exocytosis [65]. Among vegetables, dandelion is one of the richest source of beta-carotene (11,000 µg/100 g leaves, same as in carrots), from which vitamin A originates [66]. In the past few years, dandelion has demonstrated health benefits including anti-rheumatic, anti-carcinogenic, diuretic, laxative, hypoglycemic, and chloretic effects [67].
4. Anti-diabetic properties of dandelion and its components
The bioactive components in dandelion have demonstrated a series of anti-diabetic effects, which are due to the pharmacological actions of components such as sesquiterpene lactones, triterpenes/phytosterols (taraxasterol), phenols, flavonoids, and phenolic acids [44, 67]. Metformin is currently the first choice and most used anti-diabetic drug treatment, and was obtained originally from galegine discovered in Galega officinalis [68]. Similarly, acarbose, used as an anti-diabetic drug for the inhibition of alpha glucosidase, was discovered from a bacterium [69].
The main factor in T2D is the dysregulation of insulin secretion and insulin sensitivity that leads to increased blood sugar levels (hyperglycemia) and T2D, which can later cause the development of vascular diseases [70]. As T2D is both an epidemic phenomenon and a huge economic and social burden, many countries are becoming more reliant on anti-diabetic medicines [27, 28]. The root of dandelion contains inulin which includes fructo-oligosaccharides (FOS). FOS is a complex carbohydrate; its intake benefits bifido-bacteria, which eliminate pathogens in the gastrointestinal tract [66]. As a result of mineral absorption, FOS stimulates the immune system, and thereby suppresses abnormal cell growth. This complex carbohydrate can help to normalize blood sugar levels. According to Amin et al. (2015), it reduces hyperglycemia when used in high levels of water extract [66]. CGA has been a potential compound for preventing obesity and inflammation. It also impacts on insulin secretion and sensitivity, making it an attractive option for use as a future anti-diabetic drugs [61, 71].
4.1 Anti-hyperglycemic actions
Insulin resistance, which occurs in several key tissues such as liver, muscles, and adipose tissue, is the primary cause of hyperglycemia and a hallmark in T2D pathogenesis [72]. Another well-known mechanism that affects glucose homeostasis is oxidative stress, which is driven by auto-oxidation and protein glycation [73]. This process may cause an increase in the production of lipid peroxide which in turn decreases the anti-oxidative defense [67], thus supporting the development of β-cell dysfunction [74]. β-cell dysfunction impairs insulin secretion as a result of glucotoxicity and lipotoxicity, which negatively influences the conversion of proinsulin to insulin [75]. Studies on dandelion extracts revealed that it may stimulate the release of insulin in pancreatic β-cells, which consequently counteracts the effects of hyperglycemia [76].
Hussain et al. (2004) demonstrated that rat insulinoma cells (INS-1E cells) have insulin activity. Dried ethanolic extract (40 µg/ml) of Taraxacum officinale (TO) was given to the cells in the presence of high glucose (6.0 mM), using glibenclamide (an anti-diabetic drug) as a control. The authors found a significant insulin secretion by INS-1E cells compared to normal glucose (3.0 mM) [76].
Studies have revealed that CRA also increases glucose uptake in muscle cells due to the stimulation of insulin secretion in the pancreas [61]. Dandelion, administered as a 9.7% herbal preparation of ethanolic extract, has anti-hyperglycemic effects in non-obese diabetic mice [23]. Furthermore, CRA and TS inhibit α-glucosidase and α-amylase, preventing the digestion of complex carbohydrates such as starch, and thus further contributing to the anti-hyperglycemic effect [44]. Studies in diabetic rats have demonstrated that dandelion induces a decrease in glucose plasma concentration, which improves the insulin secretion activity of β-cells [44].
4.2 Anti-inflammatory action
Seo et al. (2005) showed that dandelion leaf extract has anti-inflammatory properties, which may protect against cholecystokinin-induced acute pancreatitis in rats [77]. Cholecystokinin is known to exert trophic effects in several species. TO has been used to cure liver and gallbladder disorders, which is attributable to its content of terpenoid and bitter sterol components such as taraxacin and taraxacerin [77]. Koo et al. (2004) found that TO induces apoptosis of human hepatoma (HepG2) cells through tumor necrosis factor α (TNF-α) and interleukin (IL) 1α secretion, implying anti-inflammatory effects within the central nervous system. TS is a component of TO, and has anti-carcinogenic and anti-tumor properties, as it was shown to cause a dose-dependent reduction in tumor cell viability by 26% [78]. Zhang et al. (2012) showed that TS inhibits nitric oxide (NO), prostaglandin E2, TNF-α, IL-1β, and IL-6 in lipopolysaccharide-induced macrophages. and prevents LPS-induced nuclear factor κB (NF-κB) translocation from the cytoplasm to the nucleus. These results emphasize the anti-inflammatory effect of TS, as it was shown to block the NF-κB pathway that regulates protein-inflammatory gene expression [79]. Similarly to other medicinal plants (e.g. Bidens pilosa L, Allium sativum, Gymnema sylvestre, Citrullus colocynthis, Trigonella foenum greacum, Momordica charantia, and Ficus benghalensis), TS may modulate β-cell dysfunction and increase insulin gene expression, which promotes insulin secretion from the granules and inhibits β-cell degradation [80, 81].
4.3 Anti-oxidative properties
Oxidative stress causes cellular damage involving constituents such as DNA, lipids, and proteins, eventually resulting in a metabolic disorder, which is the decisive process in the etiology of T2D [82]. Two percent of oxygen consumed end up as reactive oxygen species (ROS) in mammals. ROS include hydroxyl radicals, superoxide radicals, singlet oxygen, and peroxyl radicals, which contribute to several forms of human cancer and other severe diseases [83]. Recently, CGA found in the roots of dandelion has been identified as a potent anti-oxidant, which may suppress oxidative stress markers such as malondialdehyde and glutathione [84-86].
It has also been shown that glucose may generate ROS in β-cells, implying that glucose-induced oxidative stress is a mechanism of glucose toxicity [87]. The process of ROS formation involves auto-oxidation, oxidative phosphorylation, glycosylation, and glucosamine pathways [88]. Excess ROS production requires anti-oxidant defense, which is provided by dandelion extract, as is known from several studies conducted both in vitro and in vivo [89]. Flowers from dandelion are potential anti-oxidant resources, exerting their effect by way of their rich content of phenolic components including flavonoids, coumaric acid, and ascorbic acid [67]. Their leaf extracts are effective hydrogen donors, hydrogen peroxide scavengers, and reducing agents [90]. Several studies have demonstrated the anti-oxidative effect of dandelion. According to Hagymasi et al. (2000), extracts from dandelion leaf and root are hydrogen-donating, ROS formation-inhibiting, and radical-scavenging [91]. In another recent study of dandelion flower extracts, ethyl acetate fraction scavenged ROS by preventing DNA from ROS-induced damage. The prevention of oxidative stress was due to the presence of bioactive components including luteolin and luteolin 7-O-glucoside [92].
Obesity is a major aspect of metabolic syndrome which causes β-cell dysfunction. Failure of β-cells to produce adequate amounts of insulin is attributed to high levels of free fatty acids present in the plasma. This results in a decrease of glucose transportation into the muscle cells, thereby increasing levels of glucose and fat in the blood plasma, eventually causing hyperglycemia and lipid oxidation, which can be controlled by the anti-oxidative property of dandelion [93, 94]. In a study evaluating the effects of anti-oxidant on C57BL/6J mice fed a high-fat and -cholesterol diet using leafy mixed vegetable extracts (including dandelion), a significant drop in lipid peroxidation in various organs including the liver was observed [95]. This was due to the anti-oxidative activities expressed by enzymes present in the leafy extracts, including glutathione peroxidase, glutathione reductase, and superoxide dismutase [95].
4.4 Action mechanisms of dandelion in T2D
T2D impacts many biological systems that influence the proper function of lipid metabolism, glucose metabolism, and insulin regulation. Glucose is the main energy source for most organs of the body and insufficient release of insulin by the β-cells to control glucose levels leads to metabolic disorders. Therefore, a possible explanation for the effects and mechanisms of dandelion on T2D could be its interaction with factors involved in the metabolic syndrome (lipid metabolism, glucose metabolism, protein metabolism, α- and β-cells dysfunction) [96].
Edwin et al. (2008) highlighted the following mechanisms by which plant-derived compounds manifest their anti-diabetic properties:
- Inhibition of renal glucose reabsorption
- Reduction of the activity of carbohydrate enzymes (α-amylase with β-galactosidase and α-glucosidase)
- Reduction of dietary blood sugar (which stimulates hepatic glycolysis and glycogenesis)
- Inhibition of potassium channel flow
The mechanisms of plant polyphenolic components against T2D involve the stimulation of cAMP that increases exocytosis in β-cells, inhibition of insulin degradative processes, prevention of oxidative stress, regeneration of β-cells, reparation and cellular hypertrophy, and cellular proliferation in the islets of Langerhans [6, 74, 97-101]. Other active constituents existing in medicinal plants like dandelion include alkaloids, glycosides, amino acids, terpenoids, inorganic ions, steroids, carbohydrates, and galactomannan gum. These components have been shown to affect glucose uptake and metabolism both directly or indirectly [98].
4.5 Action of dandelion on digestion, glycolysis, and Krebs cycle
Carbohydrate consists of the constituents starch and sucrose, which supply more than 80% of the rapid energy required by the body. During digestion, most enzymes in the mouth and small intestines of humans, including α-amylase and α-glucosidase, facilitate the cleavage of carbohydrates to produce glucose which is absorbed through the walls of the small intestines into the blood stream [102]. To regulate glucose level, components like TS and CGA may inhibit α-carbohydrates. This inhibitory action contributes to glucose homeostasis, especially in diabetic patients [28]. The actions of CGA and TS to decrease glucose absorption could be a therapeutic approach that helps to reduce postprandial blood glucose level in T2D patients [66]. The bioactive components in dandelion are known to act at target sites of biochemical pathways, especially at sites where glucose metabolism is involved.
Mitochondria play another critical role in the onset of insulin resistance as they are the site at which the Krebs cycle and fatty acid oxidation take place. Their dysfunction may cause the accumulation of fat in muscle tissue and subsequently the decrease of adenosine triphosphate (ATP) in membrane transport [103]. Bioactive components in dandelion may be able to regulate these pathways, possibly via inhibition of certain enzymes that digest carbohydrates [98]. The pathways involved are the glycolytic cycle and Krebs cycle, in addition to other pathways involved in the release of insulin from β-cells. CGA and CRA may activate glucokinase in glycolysis, which catalyzes the phosphorylation of glucose to glucose-6-phosphate (G6P) [104, 105].
Glycolysis is a metabolic pathway in which a 6-carbon glucose molecule is oxidized to result in two pyruvic acid molecules. This glycolytic pathway mainly produces energy in tissues that remain in a low-oxygen state, e.g. those with low-oxygenated red blood cells. The catalytic reactions involve the actions of several enzymes, including phosphofructokinase, hexokinase, and pyruvate kinase. Bioactive components from medicinal plants such as dandelion are known to regulate enzymes such as hexokinase, glucokinase, and phosphofructokinase. These enzymes are involved in the processes of glycolysis and the Krebs cycle [98]. Studies have shown that plants rich in CGA can improve the functional activities of these crucial enzymes (phosphofructokinase, hexokinase, and pyruvate kinase) [106].
There are many enzymes involved in glycolysis and the citric acid cycle. Some components in plants are able to affect these enzymes. These components are also present in dandelion. Dandelion in particular has a high concentration of CGA, which directly affects glycolysis and indirectly the Krebs cycle. It is possible to speculate that other potent plant components such as TS and CRA, may also be responsible for the effects related to the treatment of T2D. More research on TS and CRA is required in this area.
4.6 Dandelion action during the insulin release mechanism
The mechanism of insulin release in β-cells is a complex process, which has to cope with the frequent fluctuations of glucose in the blood. Didier et al. (2008) have shown that CGA is an inhibitor of glucose-6-phosphatase (G6P) in rat liver, and may increase glucose transport, thereby increasing ATP production and stimulating insulin secretion [61]. Before insulin is released from the granules that are assembled within the trans-Golgi network in the cytoplasm of β-cells, two processes take place: 1) closure of the ATP-gated potassium channel and 2) activation of the voltage-gated calcium channel take place [107]. Jensen et al. (2008) proposed that insulin exocytosis is controlled by the intracellular ATP/ADP ratio of β-cells, followed by an elevation in glucose metabolism [108]. The enhanced ATP/ADP ratio induces plasma membrane depolarization by the closure of β-cells KATP-sensitive channels. The resultant influx of Ca2+ causes insulin export through fusion of a readily releasable pool containing vesicles within the plasma membrane [109].
Research has demonstrated that certain bioactive plant components, in particular CRA and CGA, can act on sulfonylurea-binding site 1 (SUR1), which contributes to the closure of the ATP-sensitive potassium channel (KATP), and eventually leads to membrane depolarization that promotes calcium ion influx [61, 81]. This mechanism is represented in Figure 2.
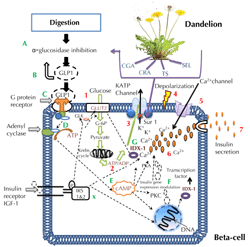 |
|
Figure 2. Potential biological mechanisms of dandelion-derived compounds to modulate cAMP and insulin secretion. The diagram specifies potential mechanisms of bioactive components from dandelion (CGA, CRA, TS, and SEL) on insulin secretion and cAMP pathway. T2D can arise from several defects, including low response in β-cells (at the level of GLUT2, SUR1, G-protein-coupled receptors, and gene expression), enzymatic action in digestion, glycolysis, and Krebs cycle. Bioactive compounds may act directly or indirectly in a series of processes, thereby modulating and regulating some T2D defects and responses. The steps of insulin secretion are shown by numbers 1-7, with steps 1 and 2 showing how ATP is produced, steps 2-5 indicating the mechanism of KATP closure and opening of the calcium ion channels, and 5-7 showing the influx of calcium ions into the β-cell triggering insulin granules to release insulin via exocytosis. A to G shows the potential mode of action of CGA, CRA, TS, and SEL at various levels of β-cell activity following the inhibition of α-glucosidase during digestion in the small intestines. A and B represent the upregulation in the release of GLP-1 secreted by intestinal L-cells, and the subsequent inhibition of glucose absorption induced by dandelion components CGA, CRA and TS. C and D indicate the stimulation of G-protein-coupled receptors and activation of adenyl cyclase, which further activates steps E and F, where cAMP activates PKA, PKC, and IDX-1, resulting in gene modulation of insulin and GLUT2, as well as an increase in calcium flow, which eventually facilitates the release of insulin. X represents the modulation of the insulin receptor by dandelion-derived compounds, which induces insulin secretion via regulation of IDX 1 factor, GLUT2, glucokinase, and endoplasmic reticulum Ca ions. |
|
Also, plant components may stimulate and increase the biosynthesis of cyclic adenosine monophosphate (cAMP), which acts as a second messenger in β-cells by inducing cholecystokinin- and glucose-stimulated insulin release [6]. Therefore, it is possible that dandelion-derived compounds prepared from TS, CGA, CRA, and other components of medicinal plants, may be able to modulate insulin gene expression, promote insulin secretion from granules, and inhibit the degradation of β-cells [81].
Finally, dandelion is very rich in ions such as calcium, which may possibly affect calcium turnover and mobilization, increasing Ca2+ plasma levels and the production of triacylglycerol (TAG), and activating isoforms of protein kinase (PKC). The effect of dandelion-derived compounds on PKA and PKC also helps to provide elevated Ca2+ levels. Most importantly, this pathway enhances the effect on insulin release [65, 110].
4.7 Mechanism of dandelion components in the regulation of β-cell gene expression
Similar to coffee, dandelion roots and flowers contain bioactive anti-oxidant components such as CGA that regulate the function of β-cells [111]. The incretin hormone glucagon-like peptide-1 (GLP-1) is secreted by intestinal L-cells as a result of the inhibition of glucose absorption by CGA [111]. Pancreatic islets consist of four cells: insulin-secreting β-cell, glucagon-secreting α-cell, somatostatin-secreting δ-cell, and polypeptide-secreting F-cell [112]. GLP-1 production, resulting from the action of CGA, helps to modulate glucose-dependent insulin secretion from the β-cell. Eventually, there is a stimulation of membrane receptors which activates cAMP [113]. GLP-1 also increases gene expression, primarily by upregulating the activities of the homeodomain islet/duodenum homeobox-1 (IDX-1), which results from the increased activity of the cAMP pathway. IDX-1 is a master regulator responsible for β-cell differentiation and function, regulation of somatostatin, and pancreatic development [114].
IDX-1 is a transcription factor, and plays an important role in insulin gene transcription, most essentially in the transcription of glucose transporter 2 (GLUT-2) and glucokinase, which initiates the glucose responsiveness in β-cells [111, 114]. Other β-cell transcription factors that regulate insulin gene expression and β-cell function include PPAR, Nkx2-2, Nkx6.1, pdx1, Beta2, Pax6, and Foxa2 [115]. Transcription factors such as the three peroxisome proliferator-activated receptors (PPAR-γ, -δ, and -α) increase the expression of genes that are responsible for carbohydrate and lipid metabolism in liver, gut, and adipose tissue [116]. Furthermore, the study of Christensen et al. (2009) revealed that components of extracts from medicinal plants such as those in dandelion stems, flowers, and leaves, activate PPAR-γ. Just like all nuclear receptors, the PPARs have a DNA-binding domain (DBD) and a ligand-binding domain (LBD). When activated they form dimers with another receptor called the retinoid-X-receptor (RXR) to be released from their co-repressor [117]. The PPAR/RXR complex binds to DNA in the nucleus through the DNA-binding domain, and causes the transcription of specific genes involved in the control of glucose and lipid metabolism [118]. Glitazones, also known as thiazolidinedione, have anti-diabetic activities, and activate PPAR-γ [119]. PPAR-γ exists in two forms (PPAR-γ-1 and PPAR-γ-2), with PPAR-γ-2 contributing to the regulation of genes and insulin sensitivity [120]. In an in-vivo experiment, Li et al. (2009) showed that CGA significantly increases the expression of hepatic mRNA through interaction with PPAR-α. It is thus possible that CGA activates PPAR-α [121]. Also, the presence of CGA may stimulate the production of GLP-1, reverse β-cell dysfunction, and thus help to treat T2D [122].
4.8 Action mechanism of dandelion in lipid metabolism
T2D results from β-cell dysfunction and other upstream metabolic disorders such as obesity and non-alcoholic fatty liver disease (NAFLD) [123]. While excessive consumption or inadequate metabolism of alcohol leads to alcoholic fatty liver disease (AFLD), consumption of excess carbohydrate or sugar leads to NAFLD, which is an abnormal lipid formation within the liver [124, 125]. This process may lead to lipogenesis from free fatty acids that accumulate through excess sugar (fructose) into fat or triglyceride in the liver, and may cause insulin resistance, impaired glucose metabolism, and inflammatory response [125].
In mice, dandelion leaf extract has been shown to reduce serum glucose, cholesterol, and triglyceride levels, possibly through the elevation of adenosine monophosphate-activated protein kinase (AMPK) in the liver, with a significant fall in lipid accumulation and improvement in insulin sensitivity [126]. Dandelion contains potential bioactive components (TS and CGA), which offer safe treatment and control of diabetes. It has several beneficial properties; it is anti-diabetic, anti-oxidative, and anti-inflammatory. In some studies, dandelion components were shown to act by inhibiting oxidative stress in liver injury, reducing high cholesterol, and reversing streptozotocin-induced diabetes [127]. Since oxidative stress is a prime marker of T2D, it is interesting that CGA (derived from dandelion) is highly anti-oxidative, able to increase the production of ROS-scavenging enzymes, malondialdehyde and glutathione, and to inhibit oxidative stress in the liver. This property of dandelion is beneficial in the scenario of fatty liver disease where lipid peroxidation can be reduced [85, 86].
Cho et al. (2010) have shown that CGA positively alters body fat in high-fat diet induced-obese mice [128]. This implies that CGA has anti-obesity properties, which can be observed through the downregulation of fatty acid and cholesterol biosynthesis, thereby reducing the risk of overexposure of β-cells to free fatty acids, and thus counteracting adiposity and insulin resistance effects [128]. There is substantial evidence that elevated concentrations of free fatty acids are toxic to β-cells [129]. The toxicity of the β-cells leads to a decrease in glucose-induced insulin secretion, impaired insulin gene expression, β-cell dysfunction, and increased cell death [15]. Prentki and Corkey (1996) hypothesized that, in the presence of physiological glucose concentrations, excessive fatty acids are readily removed by mitochondrial β-oxidation [130]. However, when fatty acids and glucose are both elevated, there is an accumulation of metabolites resulting from fatty acid esterification, which impairs β-cell function gradually [130]. It has also been demonstrated that dandelion extract is able to reduce hepatic lipid accumulation by activating the phosphorylation of AMP and AMPK, hence protecting against NAFLD and eventually against hepatic steatosis (fatty liver disease) [126].
The liver is responsible for the maintenance of blood glucose concentration during fasting and postprandial states. When the effect of insulin is lost, the liver initiates glycogenolysis, and increases hepatic glucose production [131]. An early manifestation of insulin resistance is the result of abnormalities in triglyceride storage and lipolysis in insulin-sensitive tissues [14]. There is a connection between obesity, T2D, and inflammation. Studies of TNF-α regulation in adipose tissues of mice have found a correlation between obesity and TNF-α concentration, both of which have been shown to increase the risk of insulin resistance [132]. Excess free fatty acid has been demonstrated to be toxic to hepatocytes, and to contribute to insulin resistance [133]. Coupled with the impact on digestion, carbohydrate metabolism, insulin release, and β-cell function, the effect of dandelion extracts on lipid metabolism, especially in liver and adipose tissue, may positively influence the modulation of T2D [67, 133].
4.9 Action mechanisms of dandelion in G-protein-coupled receptors and insulin function
G-protein-coupled receptors (GPCRs) are often key targets for a number of synthetic and plant-derived pharmaceuticals that are designed to improve the regulation of insulin and glucagon [134]. CRA, CGA, and TS can stimulate membrane receptors, and mediate the generation of ATP and cAMP [61]. Most of the GPCRs are involved in the regulation of islet function, and contain a variety of ions, fatty acids, and amino acids [135].
The α-subunit of the G-protein binds to guanosine 5'-diphosphate (GDP) when inactive and to guanosine triphosphate (GTP) when activated. When the GPCR is activated through binding with signaling molecules, ligands such as bioactive components from dandelion (CRA, CGA, TS) influence different cell functions [136]. Ligands bind to the GPCR, which propagates conformational changes, leading to an α-subunit exchange of phosphate between GDP and GTP, resulting from a loss of a phosphate group from the free GTP and the conversion of GDP to GTP on the receptor. As a result, subunits Gα-GTP and Gβγ dissociate from both the receptor and from each other, and regulate target proteins such as adenyl cyclase. Also, signaling from GPCR could be relayed through stimulatory Gs and inhibitory Gi, with Gi stimulating phospholipase c when activated. This cleaves phosphatidylinositol in the membrane, releasing inositol-1, 4,5- triphosphate (IP3) and TAG [137]. The cAMP is formed from the stimulation of adenyl cyclase using ATP. The actions of cAMP are known to be activated by plant components, which lead to protein phosphorylation that regulates the activity of a variety of proteins, and stimulates another enzyme called PKA [6, 138]. IP3 increases cytosolic Ca2+ originating from the endoplasmic reticulum [137]. PKA helps in the phosphorylation of different proteins and enzymes in the cytoplasm, and participates in the transcription of particular genes [139]. Eventually, this process facilitates the transport of glucose into the cells and tissues, mediating the signaling of insulin exocytosis [6]. Thus, some components of dandelion (CRA, TS, and CGA) could be applied to interact with G-protein-coupled receptors, and thereby to stimulate several activities in the β-cell, which may lead to insulin secretion, regulate hyperglycemia, and help in the prevention and treatment of T2D [140].
4.10 Glucose transportation and action of dandelion
Insulin stimulates skeletal glucose uptake by increasing GLUT4 translocation from intracellular storage vesicles to the plasma membrane and transverse tubules. Didier et al. (2008) showed that CGA stimulates glucose transport in muscle cells and insulin secretion in β-cells. Furthermore, insulin initiates activity in muscles, leading to the binding of insulin receptors followed by autophosphorylation. This process further induces a series of phosphorylations and protein-to-protein interactions mediating insulin signaling [141]. According to Mc Culloch et al. (2011), the influx of glucose is regulated by specific insulin independent GLUT membrane transporter proteins (GLUT1 in human and GLUT2 in rodent β-cells). The Michaelis constant (Km) of GLUT1 and GLUT2 is very high, indicating that they are active only at high extracellular glucose. GLUT 1, GLUT3, and GLUT4 are high-affinity-binding proteins, while GLUT2 is a low-affinity-binding protein. CGA is known to delay glucose transportation in the gut, and to initiate changes in the concentration of plasma GLP-1 and GIP. These properties may help to control T2D by reducing the transport of excess glucose by GLUT4 into the muscle cells and by stimulating the modulation of insulin gene expression [111, 142]. When mammalian cell types are exposed to metabolic stress, e.g. inhibition of oxidative phosphorylation and osmotic stress, there is an increase in glucose uptake by GLUT1 mediated by AMPK [143].
Finally, CRA is also an inhibitor of glucose-6-phosphatase (Glc-6-pase), which is involved in catalyzing glycogenolysis and gluconeogenesis in the liver. CRA binds the Glc-6-P transporter (T1) and 2-hydroxy-5-nitrobenzaldehyde (HNB), which inhibits the interaction between T1 and the phosphatase transporter. This is useful in the regulation of abnormally high rates of glucose production in T2D [142]. GLUT4 mediates insulin-stimulated glucose uptake by skeletal muscles, heart, and white and brown adipose tissue, and thus increases glucose admission into the muscles under hyperinsulinemic conditions [143]. The effect of CRA on glucose uptake in muscles may be exerted through the activation of AMPK which increases insulin sensitivity [61].
4.11 Action mechanism of dandelion in glucose uptake by muscle cells
The work by Touch (2008) demonstrated that CRA and CGA significantly increase insulin-induced glucose uptake in L6 cells (cultured muscle cells) because of the activation of the AMPK pathway that translocates GLUT4 to transport glucose into muscle cells [61]. The muscle cell membrane has insulin receptors that consist of two α and β subunits. The α subunits are external within the cell membrane and contain the insulin-binding sites, while the β subunits are internal and consist of tyrosine domains [136]. Once the α subunits are bound to insulin, the β subunits become phosphorylated, activating the tyrosine kinase enzyme that initiates insulin receptor substrate (IRS-1) and downstream pathways such as phosphatidylinositol 3-kinase (PI3K), AMPK, and mitogen-activated protein kinase (MAPK) [144]. This process increases the translocation of GLUT4 to the cell membrane, which further transports glucose into the muscle cell membrane.
Muscle cells and adipose tissues also contain PPARs [116]. The function of dandelion-derived compounds may significantly activate PPARs [120]. This activation improves muscle insulin sensitivity, which stimulates the GLUT4 receptor, and initiates the release of GLUT4 responsible for insulin-stimulated glucose uptake in muscle and adipose tissue [145, 146]. According to Kanaujia et al. (2010), the glucose uptake effect of compounds prepared from Capparis moonii is associated with significant insulin receptor (IR) and insulin receptor substrate 1 (IRS-1) phosphorylation, GLUT4 and PI3-kinase mRNA expression. This suggests that the components of dandelion may modulate the phosphorylation of IRS-1, PI3K, and GLUT4 mRNA expression, and may thus beneficially impact glucose uptake in muscle cells [147].
5. Toxicity of dandelion
As a result of the absence of toxins and alkaloids in its constituents, dandelion has low toxicity [77]. Studies on rabbits, mice, and rats, with rabbits treated orally with dried dandelion plant (3 to 6 g/kg body), and mice treated with dandelion ethanoic extracts, showed no significant or visible signs of toxicity [148]. However, allergic reactions are possible, and may occur in sensitive persons. Taraxinic acid and sesquiterpene lactone have been found to be the most allergic components in dandelion, causing allergic contact dermatitis and reactions in sensitive individuals [149]. In a study evaluating dandelion root extract efficacy in drug-resistant human melanoma cells, the root extract was revealed to be non-toxic to normal human cells (while it was toxic to human melanoma cells) [150].
Renowned physicians, the European commission, and the British Herbal Pharmacopoeia recommended the following range of doses for dandelion:
- Fresh leaves 4-10 g daily
- Dried leaves 4-10 g daily
- 2-5 ml of leaf tincture, three times a day
- Fresh leaf juice, 1 teaspoon twice daily,
- Fluid extract 1-2 teaspoon daily
- Fresh roots 2-8 g daily
- Dried powder extract 250-1000 mg four times a day [151]
Although these dose recommendations are rudimentary, they empower populations by providing alternative, accessible, and affordable treatments when conventional medicine is unattainable. Nonetheless, dandelion-derived compounds show great promise in treating T2D. Further research is warranted, especially regarding the isolation of its bioactive components, investigation and standardization of the pharmacokinetic and pharmacodynamic profiles of dandelion-derived compounds as pharmaceutical treatments and alternative medicines, and the comprehensive investigation of its effects through in-vitro, in-vivo, and clinical research.
6. Conclusions
Plant-based medicine is very useful in the management of T2D. Therapies originating from western medicine are costly and have poor availability for the majority of the global population, especially those in less developed countries [5]. Dandelion has been considered a key anti-diabetic plant because of its anti-hyperglycemic, anti-oxidative, and anti-inflammatory properties [44]. This is due to the various bioactive components present in dandelion, including polyphenolics, sesquiterpenes, triterpenes, and phytosterols. The most important and comprehensively studied bioactive components of dandelion are chlorogenic acid (CGA), chicory acid (CRA), taraxasterol (TS), and sesquiterpene lactones (SEL). These components possess great potential as anti-diabetic pharmaceuticals and nutraceuticals for regulating diabetes. They also have potential for use in anti-diabetic functional food. New, unpublished data obtained in our department indicate that TS may be the most potent regulator of T2D [33, 152].
Some research has been carried out to screen different plants for their anti-diabetic effects. Further research may make it possible to develop more affordable and more effective diabetes treatments derived from medical plants [101]. However, some work has already been done in vitro and in vivo to verify the effects and mechanisms of action of bioactive components of dandelion. In-vitro studies on clonal β-cell lines (INS-1E), α-cell lines, and human skeletal cell lines help to identify the specific bioactive components of the plant and the relevant mechanisms of action in diabetes. Therefore, new knowledge for improving diabetes treatment can be generated from this research, which is certainly able to reduce the impact of T2D on the world’s social and economic burden.
Nevertheless, there is a need for more research on the effects of dandelion components in human clinical trials. Studies using human diabetic patients would easily determine the potency and viability of dandelion components for use against T2D. More research is also necessary on the bioavailability and metabolism of these components in humans. Research in this area would pave the way for the further development of dandelion-derived compounds as drugs and provide more comprehensive information to those who are in need of treatment which is not available currently.
Disclosures: The authors reported no conflict of interests.
References
- Whiting DR, Guariguata L, Weil C, Shaw J. IDF diabetes atlas: global estimates of the prevalence of diabetes for 2011 and 2030. Diabetes Res Clin Pract 2011. 94(3):311-210. [DOD] [CrossRef]
- World Health Organization. WHO traditional medicine strategy. Geneva, World Health Organization, 2002. [DOD]
- Kalny P, Fijałek Z, Daszczuk A, Ostapczuk P. Determination of selected microelements in polish herbs and their infusions. Sci Total Environ 2007. 381(1-3):99-104. [DOD] [CrossRef]
- Guariguata L, Whiting DR, Hambleton I, Beagley J, Linnenkamp U, Shaw JE. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res Clin Pract 2014. 103(2):137-149. [DOD] [CrossRef]
- Din N. Inventory and identification of plants used in the treatment of diabetes in Douala Town (Cameroon). European J Med Plants 2011. 1(3):60-73. [DOD] [CrossRef]
- Marles RJ, Farnsworth NR. Antidiabetic plants and their active constituents. Phytomedicine 1995. 2(2):137-189. [DOD] [CrossRef]
- Mutalik S, Sulochana B, Chetana M, Udupa N, Uma Devi P. Preliminary studies on acute and subacute toxicity of an antidiabetic herbal preparation, Dianex. Indian J Exp Biol 2003. 41(4):316-320. [DOD]
- Kemper KJ. Dandelion (Taraxacum Officinalis), The Longwood herbal task force. 1999. http://longwoodherbal.org/dandelion/dandelion.pdf. [DOD]
- Malviya N, Jain S, Malviya S. Antidiabetic potential of medicinal plants. Acta Pol Pharm 2010. 67(2):113-118. [DOD]
- Fabricant DS, Farnsworth NR. The value of plants used in traditional medicine for drug discovery. Environ Health Perspect 2001. 109(3):69-75. [DOD] [CrossRef]
- Abdul-Ghani MA, Jenkinson CP, Richardson DK, Tripathy D, DeFronzo RA. Insulin secretion and action in subjects with impaired fasting glucose and impaired glucose tolerance: results from the Veterans Administration Genetic Epidemiology Study. Diabetes 2006. 55(5):1430-1435. [DOD] [CrossRef]
- Scheen AJ. Pathophysiology of type 2 diabetes. Acta Clin Belg 2003. 58(6):335-341. [DOD] [CrossRef]
- Kahn BB. Type 2 diabetes: when insulin secretion fails to compensate for insulin resistance. Cell 1998. 92(5):593-596. [DOD] [CrossRef]
- Lewis GF, Carpentier A, Adeli K, Giacca A. Disordered fat storage and mobilization in the pathogenesis of insulin resistance and type 2 diabetes. Endocr Rev 2014. 23(10):201-229. [DOD]
- McGarry JD, Dobbins RL. Fatty acids, lipotoxicity and insulin secretion. Diabetologia 1999. 42(2):128-138. [DOD] [CrossRef]
- Tateya S, Kim F, Tamori Y. Recent advances in obesity-induced inflammation and insulin resistance. Front Endocrinol (Lausanne) 2013. 4:93-94. [DOD]
- Etgen GJ, Wilson CM, Jensen J, Cushman SW, Ivy JL. Glucose transport and cell surface GLUT-4 protein in skeletal muscle of the obese Zucker rat. Am J Physiol 1996. 271(2 Pt 1):294-301. [DOD]
- Honek A, Martinkova Z, Saska P. Effect of size, taxonomic affiliation and geographic origin of dandelion (Taraxacum agg.) seeds on predation by ground beetles (Carabidae, Coleoptera). Basic Appl Ecol 2011. 12(1):89-96. [DOD] [CrossRef]
- Brock MT. The potential for genetic assimilation of a native dandelion species, Taraxacum ceratophorum (Asteraceae), by the exotic congener T. officinale. Am J Bot 2004. 91(5):656-663. [DOD] [CrossRef]
- Honek A, Martinkova Z, Saska P, Koprdova S. Role of post-dispersal seed and seedling predation in establishment of dandelion (Taraxacum agg.) plants. Agric Ecosyst Environ 2009. 134(2):126-135. [DOD] [CrossRef]
- Mingarro DM, Plaza A, Galan A, Vicente JA, Martinez MP, Acero N. The effect of five Taraxacum species on in vitro and in vivo antioxidant and antiproliferative activity. Food Funct 2015. 6(8):2787-2793. [DOD] [CrossRef]
- Kirschner J, Zaveska Drabkova L, Stepanek J, Uhlemann I. Towards a better understanding of the Taraxacum evolution (Compositae-Cichorieae) on the basis of nrDNA of sexually reproducing species. Plant Syst Evol 2014. 301(4):1135-1156. [DOD] [CrossRef]
- Petlevski R, Hadzija M, Slijepcevic M, Juretic D, Petrik J. Glutathione S-transferases and malondialdehyde in the liver of NOD mice on short-term treatment with plant mixture extract P-9801091. Phytother Res 2003. 17(4):311-314. [DOD] [CrossRef]
- Beguin P, Nagashima K, Gonoi T, Shibasaki T, Takahashi K, Kashima Y, Ozaki N, Geering K, Iwanaga T, Seino S. Regulation of Ca2+ channel expression at the cell surface by the small G-protein kir/Gem. Nature 2001. 411(6838):701-706. [DOD] [CrossRef]
- Lee S. Isolation and Identification of Phytochemical Constituents from Taraxacum coreanum. J Korean Soc Appl Biol Chem 2011. 54(1):73-78. [DOD]
- Escudero NL, de Arellano ML, Fernandez S, Albarracin G, Mucciarelli S. Taraxacum officinale as a food source. Plant Foods Hum Nutr 2003. 58(3):1-10. [DOD] [CrossRef]
- Alarcon-Aguilara F, Roman-Ramos R, Perez-Gutierrez S, Aguilar-Contreras A, Contreras-Weber C, Flores-Saenz J. Study of the anti-hyperglycemic effect of plants used as antidiabetics. J Ethnopharmacol 1998. 61(2):101-110. [DOD] [CrossRef]
- Onal S, Timur S, Okutucu B, Zihnioglu F. Inhibition of alpha-glucosidase by aqueous extracts of some potent antidiabetic medicinal herbs. Prep Biochem Biotechnol 2005. 35(1):29-36. [DOD] [CrossRef]
- Khoo HE, Prasad KN, Kong KW, Jiang Y, Ismail A. Carotenoids and their isomers: color pigments in fruits and vegetables. Molecules 2011. 16(2):1710-1738. [DOD] [CrossRef]
- Schütz K, Kammerer DR, Carle R, Schieber A. Characterization of phenolic acids and flavonoids in dandelion (Taraxacum officinale WEB. ex WIGG.) root and herb by high-performance liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun Mass Spectrom 2005. 19(2):179-186. [DOD] [CrossRef]
- Kenny O, Smyth TJ, Hewage CM, Brunton NP. Quantitative UPLC-MS/MS analysis of chlorogenic acid derivatives in antioxidant fractionates from dandelion (Taraxacum officinale) root. Int J Food Sci Technol 2015. 50(3):766-773. [DOD] [CrossRef]
- Fraisse D, Felgines C, Texier O, Lamaison J. Caffeoyl derivatives: major antioxidant compounds of some wild herbs of the Asteraceae family. Food Nutr Sci 2011. 2011:181-192. [DOD] [CrossRef]
- Williams C, Goldstone F, Greenham J. Flavonoids, cinnamic acids and coumarins from the different tissues and medicinal preparations of Taraxacum officinale. Phytochemistry 1996. 42(1):121-127. [DOD] [CrossRef]
- Westerman L, Roddick JG. Annual variation in sterol levels in leaves of Taraxacum officinale Weber. Plant Physiol 1981. 68(4):872-875. [DOD] [CrossRef]
- Roberfroid MB. Concepts in functional foods: the case of inulin and oligofructose. J Nutr 1999. 129(7 Suppl):1398S-1401S. [DOD]
- Choi UK, Lee OH, Yim JH, Cho CW, Rhee YK, Lim SI, Kim YC. Hypolipidemic and antioxidant effects of dandelion (Taraxacum officinale) root and leaf on cholesterol-fed rabbits. Int J Mol Sci 2010. 11(1):67-78. [DOD] [CrossRef]
- Arpadjan S, Celik G, Taskesen S, Gücer S. Arsenic, cadmium and lead in medicinal herbs and their fractionation. Food Chem Toxicol 2008. 46(8):2871-2875. [DOD] [CrossRef]
- Sharma K, Zafar R. Simultaneous estimation of taraxerol and taraxasterol in root callus cultures of Taraxacum officinale Weber. Int J Pharmacogn Phytochem Res 2014. 6(3):540-546. [DOD]
- Picman AK. Biological activities of sesquiterpene lactones. Biochem Syst Ecol 1986. 14(3):255-281. [DOD] [CrossRef]
- Yu F, Utsumi R. Diversity, regulation, and genetic manipulation of plant mono- and sesquiterpenoid biosynthesis. Cell Mol Life Sci 2009. 66(18):3043-3052. [DOD] [CrossRef]
- Kisiel W, Barszcz B. Further sesquiterpenoids and phenolics from Taraxacum officinale. Fitoterapia 2000. 71(3):269-273. [DOD] [CrossRef]
- Soares S, Kohl S, Thalmann S, Mateus N, Meyerhof W, De Freitas V. Different phenolic compounds activate distinct human bitter taste receptors. J Agric Food Chem 2013. 61(7):1525-1533. [DOD] [CrossRef]
- Bufe B, Hofmann T, Krautwurst D, Raguse JD, Meyerhof W. The human TAS2R16 receptor mediates bitter taste in response to beta-glucopyranosides. Nat Genet 2002. 32(3):397-401. [DOD] [CrossRef]
- Schütz K, Carle R, Schieber A. Taraxacum - a review on its phytochemical and pharmacological profile. J Ethnopharmacol 2006. 107(3):313-323. [DOD] [CrossRef]
- Lyss G. The Anti-inflammatory sesquiterpene lactone helenalin inhibits the transcription factor NF-kappa B by directly targeting p65. J Biol Chem 1998. 273(50):33508-33516. [DOD] [CrossRef]
- Schmidt TJ, Thomas J. Structure-activity relationships of sesquiterpene lactones. Stud Nat Prod Chem 2006. 33:309-392. [DOD] [CrossRef]
- Graziani G, Ferracane R, Sambo P, Santagata S, Nicoletto C, Fogliano V. Profiling chicory sesquiterpene lactones by high resolution mass spectrometry. Food Res Int 2015. 67:193-198. [DOD] [CrossRef]
- Chadwick M, Trewin H, Gawthrop F, Wagstaff C. Sesquiterpenoids lactones: benefits to plants and people. Int J Mol Sci 2013. 14(6):12780-127805. [DOD] [CrossRef]
- Koh YJ, Cha DS, Ko JS, Park HJ, Choi HD. Anti-inflammatory effect of Taraxacum officinale leaves on lipopolysaccharide-induced inflammatory responses in RAW 264.7 cells. J Med Food 2010. 13(4):870-878. [DOD] [CrossRef]
- Xu R, Fazio GC, Matsuda SP. On the origins of triterpenoid skeletal diversity. Phytochemistry 2004. 65(3):261-291. [DOD] [CrossRef]
- Ovesna Z, Vachalkova A, Horvathova K. Taraxasterol and beta-sitosterol: new naturally compounds with chemoprotective/chemopreventive effects. Neoplasma 2004. 51(6):407-414. [DOD]
- Akihisa T, Yasukawa K, Oinuma H, Kasahara Y, Yamanouchi S, Takido M, Kumaki K, Tamura T. Triterpene alcohols from the flowers of compositae and their anti-inflammatory effects. Phytochemistry 1996. 43(6):1255-1260. [DOD] [CrossRef]
- Abidi S. Chromatographic analysis of plant sterols in foods and vegetable oils. J Chromatogr A 2001. 935(1-2):173-201. [DOD] [CrossRef]
- Villarreal ML, Alvarez L, Alonso D, Navarro V, Garcia P, Delgado G. Cytotoxic and antimicrobial screening of selected terpenoids from Asteraceae species. J Ethnopharmacol 1994. 42(1):25-29. [DOD] [CrossRef]
- Kenny O, Smyth TJ, Hewage CM, Brunton NP. Antioxidant properties and quantitative UPLC-MS/MS analysis of phenolic compounds in dandelion (Taraxacum officinale) root extracts. Free Radicals Antioxidants 2014. 4(1):55-61. [DOD] [CrossRef]
- Davaatseren M, Hur HJ, Yang HJ, Hwang JT, Park JH, Kim HJ, Kim MS, Kim MJ, Kwon DY, Sung MJ. Dandelion leaf extract protects against liver injury induced by methionine- and choline-deficient diet in mice. J Med Food 2013. 16(1):26-33. [DOD] [CrossRef]
- Colle D, Arantes LP, Gubert P, da Luz SCA, Athayde ML, Teixeira Rocha JB, Soares FA. Antioxidant properties of Taraxacum officinale leaf extract are involved in the protective effect against hepatoxicity induced by acetaminophen in mice. J Med Food 2012. 15(6):549-556. [DOD] [CrossRef]
- Chkhikvishvili ID, Kharebava GI. Chicoric and chlorogenic acids in various plants growing in Georgia. Prikl Biokhim Mikrobiol 2001. 37(2):214-217. [DOD]
- Kang J, Liu Y, Xie MX, Li S, Jiang M, Wang YD. Interactions of human serum albumin with chlorogenic acid and ferulic acid. Biochim Biophys Acta 2004. 1674(2):205-214. [DOD] [CrossRef]
- Nakatani N, Kayano S, Kikuzaki H, Sumino K, Katagiri K, Mitani T. Identification, Quantitative Determination, and Antioxidative Activities of Chlorogenic Acid Isomers in Prune (Prunus d omestica L.). J Agric Food Chem 2000. 48(11):5512-5516. [DOD] [CrossRef]
- Tousch D, Lajoix AD, Hosy E, Azay-Milhau J, Ferrare K, Jahannault C, Cros G, Petit P. Chicoric acid, a new compound able to enhance insulin release and glucose uptake. Biochem Biophys Res Commun 2008. 377(1):131-145. [DOD] [CrossRef]
- Kim M. The water-soluble extract of chicory reduces cholesterol uptake in gut-perfused rats. Nutr Res 2000. 20(7):1017-1026. [DOD] [CrossRef]
- Kim M, Shin HK. The water-soluble extract of chicory influences serum and liver lipid concentrations, cecal short-chain fatty acid concentrations and fecal lipid excretion in rats. J Nutr 1998. 128(10):1731-1736. [DOD]
- Ata S, Farooq F, Javed S. Elemental profile of 24 common medicinal plants of Pakistan and its direct link with traditional uses. J Med Plants Res 2011. 5(26):6164-6168. [DOD]
- Komatsu M, Schermerhorn T, Noda M, Straub SG, Aizawa T, Sharp GW. Augmentation of insulin release by glucose in the absence of extracellular Ca2+: new insights into stimulus-secretion coupling. Diabetes 1997. 46(12):1928-1938. [DOD] [CrossRef]
- Mir MA, Sawhney SS, Jassal MM. In-vitro antidiabetic studies of various extracts of Taraxacum officinale. Pharma Innov 2015. 4(1):61-66. [DOD]
- Gonzalez-Castejon M, Visioli F, Rodriguez-Casado A. Diverse biological activities of dandelion. Nutr Rev 2012. 70(9):534-547. [DOD] [CrossRef]
- Bailey C, Day C. Metformin: its botanical background. Pract Diabetes Int 2004. 21(3):115-117. [DOD] [CrossRef]
- Brunkhorst C, Schneider E. Characterization of maltose and maltotriose transport in the acarbose-producing bacterium Actinoplanes sp. Res Microbiol 2005. 156(8):851-857. [DOD] [CrossRef]
- Resnick HE, Howard BV. Diabetes and cardiovascular disease. Annu Rev Med 2002. 53:245-267. [DOD] [CrossRef]
- Xiao H, Xie G, Wang J, Hou X, Wang X, Wu W, Liu X. Chicoric acid prevents obesity by attenuating hepatic steatosis, inflammation and oxidative stress in high-fat diet-fed mice. Food Res Int 2013. 54(1):345-353. [DOD] [CrossRef]
- Hamden K, Carreau S, Boujbiha MA, Lajmi S, Aloulou D, Kchaou D, Elfeki A. Hyperglycaemia, stress oxidant, liver dysfunction and histological changes in diabetic male rat pancreas and liver: protective effect of 17 beta-estradiol. Steroids 2008. 73(5):495-501. [DOD] [CrossRef]
- Giugliano D, Ceriello A, Paolisso G. Oxidative stress and diabetic vascular complications. Diabetes Care 1996. 19(3):257-267. [DOD] [CrossRef]
- Kaneto H, Katakami N, Kawamori D, Miyatsuka T, Sakamoto K, Matsuoka TA, Matsuhisa M, Yamasaki Y. Involvement of oxidative stress in the pathogenesis of diabetes. Antioxid Redox Signal 2007. 9(3):355-366. [DOD] [CrossRef]
- Cheng DM, Pogrebnyak N, Kuhn P, Krueger CG, Johnson WD, Raskin I. Development and phytochemical characterization of high polyphenol red lettuce with anti-diabetic properties. Plos One 2014. 9(3):934-411. [DOD]
- Hussain Z, Waheed A, Qureshi RA, Burdi DK, Verspohl EJ, Khan N, Hasan M. The effect of medicinal plants of Islamabad and Murree region of Pakistan on insulin secretion from INS-1 cells. Phytother Res 2004. 18(1):73-77. [DOD] [CrossRef]
- Seo S, Koo H, An H, Kwon K, Lim B, Seo E, Ryu D, Moon G, Kim H, Kim H, Hong SH. Taraxacum officinale protects against cholecystokinin-induced acute pancreatitis in rats. World J Gastroenterol 2005. 11(4):597-599. [DOD] [CrossRef]
- Koo HN, Hong SH, Song BK, Kim CH, Yoo YH, Kim HM. Taraxacum officinale induces cytotoxicity through TNF-alpha and IL-1alpha secretion in Hep G2 cells. Life Sci 2004. 74(9):1149-1157. [DOD] [CrossRef]
- Zhang X, Xiong H, Liu L. Effects of taraxasterol on inflammatory responses in lipopolysaccharide-induced RAW 264.7 macrophages. J Ethnopharmacol 2012. 141(1):206-211. [DOD] [CrossRef]
- Patel DK, Prasad SK, Kumar R, Hemalatha S. An overview on antidiabetic medicinal plants having insulin mimetic property. Asian Pac J Trop Biomed 2012. 2(4):320-330. [DOD] [CrossRef]
- Jones PM, Persaud SJ. Protein kinases, protein phosphorylation, and the regulation of insulin secretion from pancreatic beta-cells. Endocr Rev 1998. 19(4):429-461. [DOD]
- Yakes FM, Van Houten B. Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells following oxidative stress. Proc Natl Acad Sci U S A 1997. 94(2):514-519. [DOD] [CrossRef]
- Hu C, Kitts DD. Dandelion (Taraxacum officinale) flower extract suppresses both reactive oxygen species and nitric oxide and prevents lipid oxidation in vitro. Phytomedicine 2005. 12(8):588-597. [DOD] [CrossRef]
- Leung AY. Encyclopedia of common natural ingredients: used in food, drugs and cosmetics. John Wiley, New York, 1996, pp. 205-207. [DOD]
- Ceballos-Picot I, Witko-Sarsat V, Merad-Boudia M, Nguyen AT, Thevenin M, Jaudon MC, Zingraff J, Verger C, Jingers P, Descamps-Latscha B. Glutathione antioxidant system as a marker of oxidative stress in chronic renal failure. Free Radic Biol Med 1996. 21(6):845-853. [DOD] [CrossRef]
- Del Rio D, Stewart AJ, Pellegrini N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr Metab Cardiovasc Dis 2005. 15(4):316-328. [DOD] [CrossRef]
- Robertson R, Zhou H, Zhang T, Harmon JS. Chronic oxidative stress as a mechanism for glucose toxicity of the beta cell in type 2 diabetes. Cell Biochem Biophys 2007. 48(2-3):139-146. [DOD] [CrossRef]
- Robertson RP, Harmon J, Tran PO, Poitout V. Beta-cell glucose toxicity, lipotoxicity, and chronic oxidative stress in type 2 diabetes. Diabetes 2004. 53(1):119-124. [DOD] [CrossRef]
- You Y, Yoo S, Yoon HG, Park J, Lee YH, Kim S, Oh KT, Lee J, Cho HY, Jun W. In vitro and in vivo hepatoprotective effects of the aqueous extract from Taraxacum officinale (dandelion) root against alcohol-induced oxidative stress. Food Chem Toxicol 2010. 48(6):1632-1637. [DOD] [CrossRef]
- Czinner E, Hagymasi K, Blazovics A, Kery A, Szoke E, Lemberkovics E. The in vitro effect of Helichrysi flos on microsomal lipid peroxidation. J Ethnopharmacol 2001. 77(1):31-35. [DOD] [CrossRef]
- Hagymasi K, Blazovics A, Feher J, Lugasi A, Kristo ST, Kery A. The in vitro effect of dandelions antioxidants on microsomal lipid peroxidation. Phytother Res 2000. 14(1):43-44. [DOD] [CrossRef]
- Hu C, Kitts DD. Dandelion (Taraxacum officinale) flower extract suppresses both reactive oxygen species and nitric oxide and prevents lipid oxidation in vitro. Phytomedicine 2005. 12(8):588-597. [DOD] [CrossRef]
- Tfayli H, Bacha F, Gungor N, Arslanian S. Phenotypic type 2 diabetes in obese youth: insulin sensitivity and secretion in islet cell antibody-negative versus -positive patients. Diabetes 2009. 58(3):738-744. [DOD] [CrossRef]
- Boden G. Role of fatty acids in the pathogenesis of insulin resistance and NIDDM. Diabetes 1997. 46(1):3-10. [DOD] [CrossRef]
- Kim MY, Kim MH, Son C, Yook H, Kim JH, Cho Y, Chun H, Kim MR. Leafy vegetable mix supplementation improves lipid profiles and antioxidant status in C57BL/6J mice fed a high fat and high cholesterol diet. J Med Food 2009. 12(4):877-884. [DOD] [CrossRef]
- Grundy SM, Brewer HB, Cleeman JI, Smith SC, Lenfant C. Definition of metabolic syndrome: Report of the National Heart, Lung, and Blood Institute/American Heart Association conference on scientific issues related to definition. Circulation 2004. 109(3):433-438. [DOD] [CrossRef]
- Eddouks M, Maghrani M, Lemhadri A, Ouahidi ML, Jouad H. Ethnopharmacological survey of medicinal plants used for the treatment of diabetes mellitus, hypertension and cardiac diseases in the south-east region of Morocco (Tafilalet). J Ethnopharmacol 2002. 82(2-3):97-103. [DOD] [CrossRef]
- Prabhakar PK, Doble M. A target based therapeutic approach towards diabetes mellitus using medicinal plants. Curr Diabetes Rev 2008. 4(4):291-308. [DOD] [CrossRef]
- Mukherjee PK, Maiti K, Mukherjee K, Houghton PJ. Leads from Indian medicinal plants with hypoglycemic potentials. J Ethnopharmacol 2006. 106(1):1-28. [DOD] [CrossRef]
- Bnouham M, Ziyyat A, Mekhfi H, Tahri A, Legssyer A. Medicinal plants with potential antidiabetic activity - a review of ten years of herbal medicine research (1990-2000). Int J Diabetes Metab 2006. 14(1):1-25. [DOD]
- Jarald E, Joshi SB, Jain DC. Diabetes and herbal medicines. Iran J Pharmacol Ther 2008. 7(1):97-106. [DOD]
- Kumar S, Narwal S, Kumar V, Prakash O. Alpha-glucosidase inhibitors from plants: a natural approach to treat diabetes. Pharmacogn Rev 2011. 5(9):19-29. [DOD] [CrossRef]
- Perk J, De Backer G, Gohlke H, Graham I, Reiner Z, Verschuren WM, Albus C, Benlian P, Boysen G, Cifkova R, et al. European guidelines on cardiovascular disease prevention in clinical practice (version 2012). The fifth joint task force of the European Society of Cardiology and other societies on cardiovascular disease prevention in clinical practice. G Ital Cardiol (Rome) 2013. 14(5):328-392. [DOD]
- Coman C, Rugina OD, Socaciu C. Plants and natural compounds with antidiabetic action. Not Bot Horti Agrobo 2012. 40(1):314-325. [DOD]
- Vessal M, Hemmati M, Vasei M. Antidiabetic effects of quercetin in streptozocin-induced diabetic rats. Comp Biochem Physiol Part C Toxicol Pharmacol 2003. 135(3):357-364. [DOD] [CrossRef]
- Prabhakar P, Doble M. A target-based therapeutic approach towards diabetes mellitus using medicinal plants. Curr Diabetes Rev 2008. 4(4):291-308. [DOD] [CrossRef]
- Bratanova-Tochkova TK, Cheng H, Daniel S, Gunawardana S, Liu YJ, Mulvaney-Musa J, Schermerhorn T, Straub SG, Yajima H, Sharp GW. Triggering and augmentation mechanisms, granule pools, and biphasic insulin secretion. Diabetes 2002. 51(1):83-90. [DOD] [CrossRef]
- Jensen J, Ruge T, Lai YC, Svensson MK, Eriksson JW. Effects of adrenaline on whole-body glucose metabolism and insulin-mediated regulation of glycogen synthase and PKB phosphorylation in human skeletal muscle. Metabolism 2011. 60(2):215-226. [DOD] [CrossRef]
- Jensen MV, Joseph JW, Ronnebaum SM, Burgess SC, Sherry AD, Newgard CB. Metabolic cycling in control of glucose-stimulated insulin secretion. Am J Physiol Endocrinol Metab 2008. 295(6):E1287-E1297. [DOD] [CrossRef]
- Chay TR. Modeling slowly bursting neurons via calcium store and voltage-independent calcium current. Neural Comput 1996. 8(5):951-978. [DOD] [CrossRef]
- McCarty MF. A chlorogenic acid-induced increase in GLP-1 production may mediate the impact of heavy coffee consumption on diabetes risk. Med Hypotheses 2005. 64(4):848-853. [DOD] [CrossRef]
- Brissova M, Fowler MJ, Nicholson WE, Chu A, Hirshberg B, Harlan DM, Powers AC. Assessment of human pancreatic islet architecture and composition by laser scanning confocal microscopy. J Histochem Cytochem 2005. 53(9):1087-1097. [DOD] [CrossRef]
- Macdonald PE, El-kholy W, Riedel MJ, Salapatek AM, Light PE, Wheeler MB. The Multiple actions of GLP-1 on the process of glucose-stimulated insulin secretion. Diabetes 2002. 51(Suppl 3):S434-S442. [DOD]
- Stoffers DA, Thomas MK, Habener JF. Homeodomain protein IDX-1. Trends Endocrinol Metab 1997. 8(4):145-151. [DOD] [CrossRef]
- Nordentoft I, Jeppesen PB, Hong J, Abudula R, Hermansen K. Isosteviol increases insulin sensitivity and changes gene expression of key insulin regulatory genes and transcription factors in islets of the diabetic KKAy mouse. Diabetes Obes Metab 2008. 10(10):939-949. [DOD] [CrossRef]
- Desvergne B, Wahli W. Peroxisome proliferator-activated receptors: nuclear control of metabolism. Endocr Rev 1999. 20(5):649-688. [DOD]
- Guri AJ, Hontecillas R, Bassaganya-Riera J. Peroxisome proliferator-activated receptors: bridging metabolic syndrome with molecular nutrition. Clin Nutr 2006. 25(6):871-885. [DOD] [CrossRef]
- Grimaldi PA. Regulatory functions of PPARbeta in metabolism: implications for the treatment of metabolic syndrome. Biochim Biophys Acta 2007. 1771(8):983-990. [DOD] [CrossRef]
- Sakamoto J, Kimura H, Moriyama S, Odaka H, Momose Y, Sugiyama Y, Sawada H. Activation of human peroxisome proliferator-activated receptor (PPAR) subtypes by pioglitazone. Biochem Biophys Res Commun 2000. 278(3):704-711. [DOD] [CrossRef]
- Christensen KB, Minet A, Svenstrup H, Grevsen K, Zhang H, Schrader E, Rimbach G, Wein S, Wolffram S, Kristiansen K, et al. Identification of plant extracts with potential antidiabetic properties: effect on human peroxisome proliferator-activated receptor (PPAR), adipocyte differentiation and insulin-stimulated glucose uptake. Phytother Res 2009. 23(9):1316-1325. [DOD] [CrossRef]
- Li SY, Chang CQ, Ma FY, Yu CL. Modulating effects of chlorogenic acid on lipids and glucose metabolism and expression of hepatic peroxisome proliferator-activated receptor-alpha in golden hamsters fed on high fat diet. Biomed Environ Sci 2009. 22(2):122-129. [DOD] [CrossRef]
- Scheen AJ. Is there a role for alpha-glucosidase inhibitors in the prevention of type 2 diabetes mellitus? Drugs 2003. 63(10):933-951. [DOD]
- Zimmet P, Alberti KG, Shaw J. Global and societal implications of the diabetes epidemic. Nature 2001. 414(6865):782-787. [DOD] [CrossRef]
- Samuel VT, Liu ZX, Qu X, Elder BD, Bilz S, Befroy D, Romanelli AJ, Shulman GI. Mechanism of hepatic insulin resistance in non-alcoholic fatty liver disease. J Biol Chem 2004. 279(31):32345-32353. [DOD] [CrossRef]
- Cornier MA, Dabelea D, Hernandez TL, Lindstrom RC, Steig AJ, Stob NR, Van Pelt RE, Wang H, Eckel RH. The metabolic syndrome. Endocr Rev 2008. 29(7):777-822. [DOD] [CrossRef]
- Davaatseren M, Hur HJ, Yang HJ, Hwang JT, Park JH, Kim HJ, Kim MJ, Kwon DY, Sung MJ. Taraxacum official (dandelion) leaf extract alleviates high-fat diet-induced nonalcoholic fatty liver. Food Chem Toxicol 2013. 58:30-36. [DOD] [CrossRef]
- Cho SY, Park JY, Park EM, Choi MS, Lee MK, Jeon SM, Jang MK, Kim MJ, Park YB. Alternation of hepatic antioxidant enzyme activities and lipid profile in streptozotocin-induced diabetic rats by supplementation of dandelion water extract. Clin Chim Acta 2002. 317(1-2):109-117. [DOD] [CrossRef]
- Cho AS, Jeon SM, Kim MJ, Yeo J, Seo KI, Choi MS, Lee MK. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem Toxicol 2010. 48(3):937-943. [DOD] [CrossRef]
- Boden G, Shulman GI. Free fatty acids in obesity and type 2 diabetes: defining their role in the development of insulin resistance and beta-cell dysfunction. Eur J Clin Invest 2002. 32(3):14-23. [DOD] [CrossRef]
- Prentki M, Corkey BE. Are the beta-cell signaling molecules malonyl-CoA and cystolic long-chain acyl-CoA implicated in multiple tissue defects of obesity and NIDDM? Diabetes 1996. 45(3):273-283. [DOD]
- Consoli A, Nurjhan N, Capani F, Gerich J. Predominant role of gluconeogenesis in increased hepatic glucose production in NIDDM. Diabetes 1989. 38(5):550-557. [DOD] [CrossRef]
- Hotamisligil GS, Shargill NS, Spiegelman BM. Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science 1993. 259(5091):87-91. [DOD] [CrossRef]
- Harris EH. Elevated liver function tests in type 2 diabetes. Clin Diabetes 2005. 23(3):115-119. [DOD] [CrossRef]
- Garrido DM, Corbett DF, Dwornik KA, Goetz AS, Littleton TR, McKeown SC, Mills WY, Smalley TL, Briscoe CP, Peat AJ. Synthesis and activity of small molecule GPR40 agonists. Bioorg Med Chem Lett 2006. 16(7):1840-1845. [DOD] [CrossRef]
- Klabunde T, Hessler G. Drug design strategies for targeting G-protein-coupled receptors. Chembiochem 2002. 3(10):928-944. [DOD] [CrossRef]
- Chang L, Chiang SH, Saltiel AR. Insulin signaling and the regulation of glucose transport. Mol Med 2005. 10(7-12):65-71. [DOD]
- Haber EP, Hirabara SM, Gomes AD, Curi R, Carpinelli AR, Carvalho CR. Palmitate modulates the early steps of insulin signalling pathway in pancreatic islets. FEBS Lett 2003. 544(1-3):185-188. [DOD] [CrossRef]
- Howe AK. Regulation of actin-based cell migration by cAMP/PKA. Biochim Biophys Acta 2004. 1692(2-3):159-174. [DOD] [CrossRef]
- Metz R, Ziff E. cAMP stimulates the C/EBP-related transcription factor rNFIL-6 to trans-locate to the nucleus and induce c-fos transcription. Genes Dev 1991. 5(10):1754-1766. [DOD] [CrossRef]
- Cheynier V, Tomas-Barberan FA, Yoshida K. Polyphenols: from plants to a variety of food and nonfood uses. J Agric Food Chem 2015. 63(35):7589-7594. [DOD] [CrossRef]
- Shepherd PR. Mechanisms regulating phosphoinositide 3-kinase signalling in insulin-sensitive tissues. Acta Physiol Scand 2005. 183(1):3-12. [DOD] [CrossRef]
- Arion WJ, Canfield WK, Ramos FC, Schindler PW, Burger HJ, Hemmerle H, Schubert G, Below P, Herling AW. Chlorogenic acid and hydroxynitrobenzaldehyde: new inhibitors of hepatic glucose 6-phosphatase. Arch Biochem Biophys 1997. 339(2):315-322. [DOD] [CrossRef]
- Pillay TS, Makgoba MW. Molecular mechanisms of insulin resistance. S Afr Med J 1991. 79(10):607-613. [DOD]
- Mlinar B, Marc J, Janez A, Pfeifer M. Molecular mechanisms of insulin resistance and associated diseases. Clin Chim Acta 2007. 375(1-2):20-35. [DOD] [CrossRef]
- Petersen KF, Shulman GI. Etiology of insulin resistance. Am J Med 2006. 119(5 Suppl 1):S10-S16. [DOD] [CrossRef]
- Kobayashi H, Mitsui T, Nomura S, Ohno Y, Kadomatsu K, Muramatsu T, Nagasaka T, Mizutani S. Expression of glucose transporter 4 in the human pancreatic islet of Langerhans. Biochem Biophys Res Commun 2004. 314(4):1121-1125. [DOD] [CrossRef]
- Kanaujia A, Duggar R, Pannakal ST, Yadav SS, Katiyar CK, Bansal V, Anand S, Sujatha S, Lakshmi BS. Insulinomimetic activity of two new gallotannins from the fruits of Capparis moonii. Bioorg Med Chem 2010. 18(11):3940-3945. [DOD] [CrossRef]
- Akhtar MS, Khan QM, Khaliq T. Effects of Portulaca oleracae (Kulfa) and Taraxacum officinale (Dhudhal) in normoglycaemic and alloxan-treated hyperglycaemic rabbits. J Pak Med Assoc 1985. 35(7):207-210. [DOD]
- Lovell CR, Rowan M. Dandelion dermatitis. Contact Dermatitis 1991. 25(3):185-188. [DOD] [CrossRef]
- Chatterjee SJ, Ovadje P, Mousa M, Hamm C, Pandey S. The efficacy of dandelion root extract in inducing apoptosis in drug-resistant human melanoma cells. Evid Based Complement Alternat Med 2011. 2011:129045. [DOD]
- Newall CA, Anderson LA, Phillipson JD. Herbal medicines. A guide for health-care professionals. Pharm Press, 1996, pp. 296-297. [DOD]
- Akashi T, Furuno T, Takahashi T, Ayabe SI. Biosynthesis of triterpenoids in cultured cells, and regenerated and wild plant organs of Taraxacum officinale. Phytochemistry 1994. 36(2):303-308. [DOD] [CrossRef]
|


)
)
)

